核小体
实验室致力于破解由组蛋白修饰酶复合物、DNA修饰酶、染色体重塑复合物等表观遗传因子所组成的表观遗传调控网络,揭示它们介导核小体修饰、组装和变构等过程的分子机制。
组蛋白甲基化修饰:甲基转移酶MLL复合物的活性调控机制
MLL家族甲基转移酶蛋白,其名称起源于该家族第一个被发现的基因—混合谱系白血病MLL1(Mixed Lineage Leukemia 1)—的基因重排在急性淋巴细胞和髓细胞白血病中高发。哺乳动物中MLL家族甲基转移酶包含MLL1,MLL2,MLL3,MLL4,SET1A和SET1B六个成员,在基因表达调控中发挥重要且非冗余的作用。MLL家族蛋白在体内和多个辅助亚基形成复合物后才具有可观的甲基转移酶活性。实验室的一系列工作揭示了WDR5、RBBP5、ASH2L、DPY30等辅助亚基调控MLL蛋白酶活的分子机制。
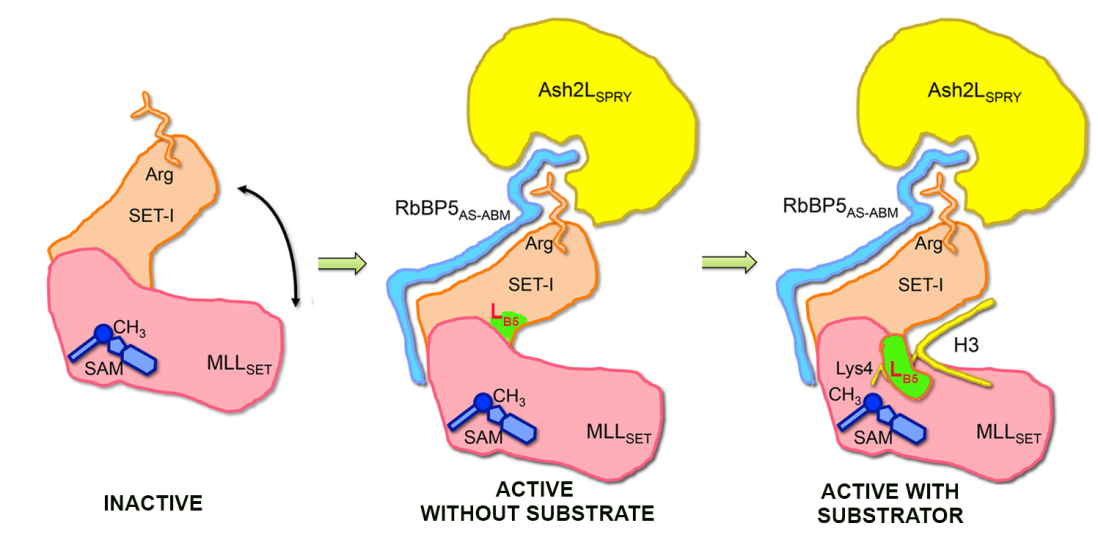
2016年,我们实验室与雷鸣实验室、李国辉实验室合作阐明了RBBP5-ASH2L异源二聚体结合和激活MLL家族蛋白的分子机制。通过解析MLL家族蛋白中一系列蛋白单体及蛋白复合物的晶体结构,辅以分子动力学模拟,证实了加入RBBP5-ASH2L能够限制SET-I结构模块的柔性,从而使灵活的MLL催化结构域固定在一种活性构象,增强了甲基转移酶活性(Nature 2016)。
https://www.nature.com/articles/nature16952
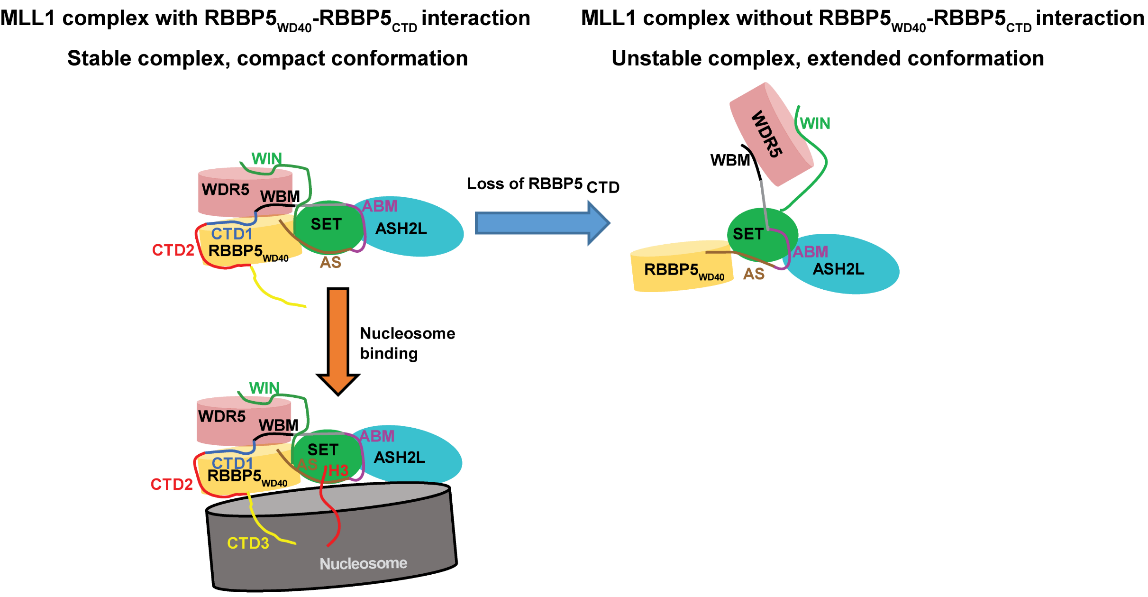
2019年,我们实验室揭示了全长RBBP5蛋白调控MLL1复合物活性的分子机制。RBBP5的C末端两个多肽模块(CTD1,2)和N端的WD40结构域存在内部的直接作用,使得MLL1复合物的构象更加紧凑,从而增强MLL1复合物的甲基转移酶活性(Nucleic Acids Research 2019)。
https://academic.oup.com/nar/article/47/19/10426/5572575
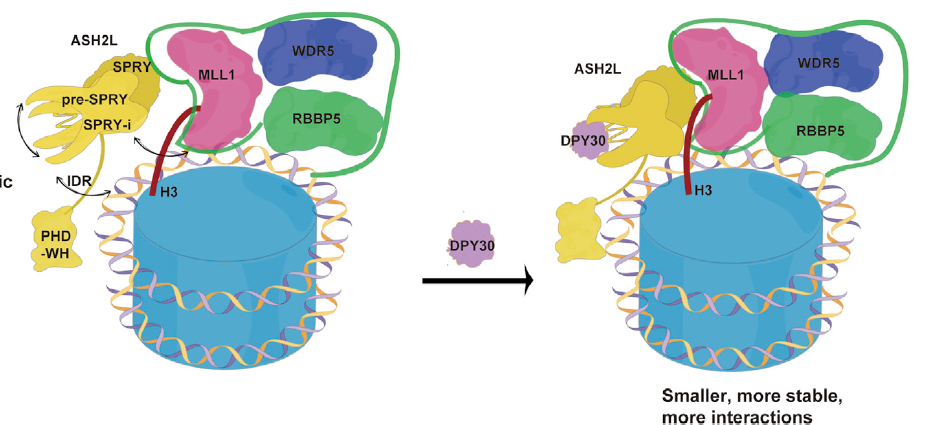
2022年,我们实验室阐明了DPY30增强MLL家族复合物酶活的双重机制。DPY30增强了ASH2L 的结构紧致性和热稳定性,一方面 DPY30 稳定后的ASH2L与RBBP5亲和力增加,促进MLL复合物组装形成一个更紧凑和更稳定的复合物;另一方面DPY30稳定后的ASH2L获得了与核小体的DNA和H3相互作用的能力,从而极大地促进了MLL复合物在核小体上的甲基转移酶活性(iScience 2022)。
https://www.cell.com/iscience/fulltext/S2589-0042(22)01220-2
2022年,我们实验室合作揭示了高度类似的MLL家族蛋白具有生成不同甲基化产物的酶活特性(即产物特异性)的分子机制。我们首先建立了定义组蛋白甲基转移酶产物特异性的定量方法,界定了MLL家族蛋白不同成员的产物特异性。进一步我们发现活性位点两个关键的酪氨酸(F/Y switch)的动态性决定了MLL家族蛋白的产物特异性: SET结构域内部不同的作用网络决定了关键酪氨酸(F/Y switch)的动态性在不同MLL成员中有较大差异,从而产生了不同的甲基化产物生成特异性(Molecular Cell 2022)。
https://doi.org/10.1016/j.molcel.2022.08.022

组蛋白酰化修饰:酰化修饰的解读和信号传递机制
2022年,我们实验室合作揭示了最近发现的组蛋白苯甲酰化修饰的建立和识别机制。苯甲酰化修饰是唯一已知的带有芳香族苯环的组蛋白修饰,与其他带有短链脂肪酸的组蛋白标记相比,苯甲酰基具有更大的分子体积和更强的疏水性。我们首次鉴定了酿酒酵母组蛋白中的30个组蛋白苯甲酰化修饰位点;其次,通过酵母遗传学筛选和体外生化实验,鉴定了苯甲酰化转移酶(SAGA)、去苯甲酰化酶(Hst2)以及苯甲酰化阅读器(Taf14、Sas5 两个蛋白的YEATS结构域和Sth1蛋白的 Bromo结构域);进而,通过结构解析和生化分析,揭示了这些蛋白催化和识别苯甲酰化修饰的分子机制(Nature Communications 2022)。

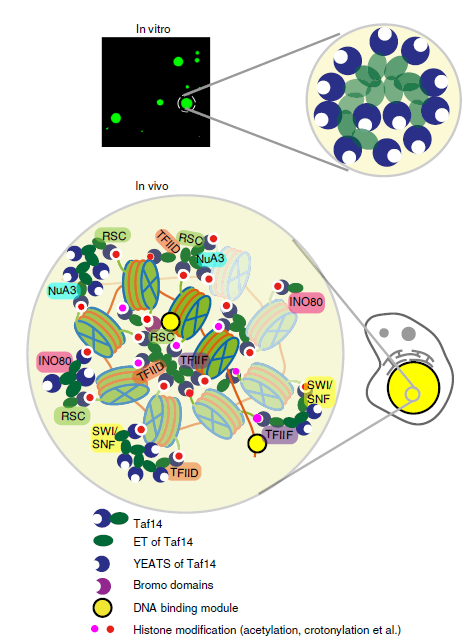
2019年,我们实验室揭示了组蛋白酰化修饰阅读器Taf14与众多转录相关复合物互作以及调控基因转录的分子机制。酿酒酵母中的YEATS结构域蛋白Taf14是一个重要的转录调控蛋白,我们揭示了Taf14与多种染色质重塑复合物(RSC,SWI / SNF,INO80)、修饰酶复合物(NuA3)、转录因子(TFIID和TFIIF)等结合的保守相互作用模式,确定了酰化修饰阅读器Taf14在染色质重塑复合物RSC中的重要功能,并建立了Taf14通过相变富集多种转录机器到特定组蛋白修饰位点来实现高效转录的工作模型(Nature Communications 2020)。
https://www.nature.com/articles/s41467-020-18021-7
异染色质
染色质高级结构的调控机制研究:以典型异染色质高级结构——端粒为范例开展异染色质调控分子的鉴定和机制解析,揭示多个重要端粒结合蛋白和表观遗传因子维持和调控异染色质高级结构、组装和定位的分子机制。
阐明了Sad1介导异染色质组装的分子机制
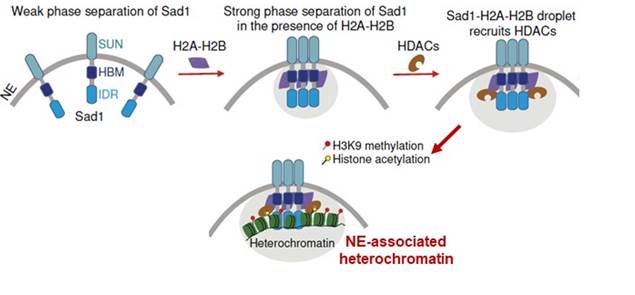
SUN家族蛋白是真核生物中高度保守的内核膜蛋白,与位于外核膜上的KASH蛋白组成LINC复合体连接核骨架与细胞骨架,在细胞核迁移、核定位以及机械力传导、染色体运动等方面发挥重要功能,但SUN家族蛋白是否参与异染色质的调控仍然未知。我们的近期工作发现裂殖酵母中的SUN家族蛋白Sad1通过和组蛋白H2A-H2B的互作,在异染色质的核膜定位和维持异染色质沉默状态中发挥重要作用。组蛋白H2A-H2B极大的促进了Sad1和一系列去乙酰化酶(Clr3,Sir2等)的相互作用,从而保证了异染色质区的低乙酰化状态。此外,H2A-H2B能促进Sad1在核膜上的相分离(phase separation),进而招募富集多种异染色质因子来介导酵母体内异染色质的形成与核膜定位(Nature Commun 2024)。
https://www.nature.com/articles/s41467-024-48418-7
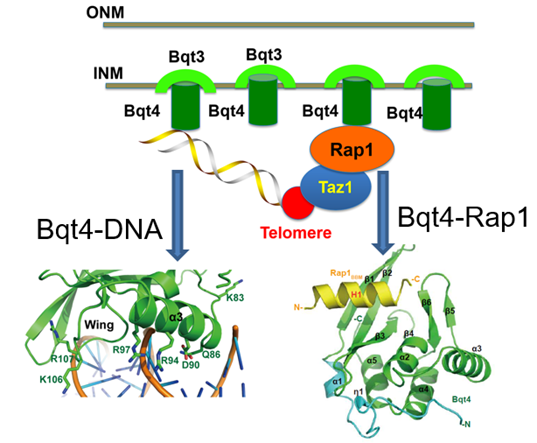
揭示了Bqt4蛋白调控端粒定位的分子机制
真核细胞端粒通常会区域性的定位于细胞核内膜周围,端粒-核膜连接对染色质稳态的维持以及细胞分裂至关重要,但端粒-核膜连接的分子细节尚不清楚。我们发现裂殖酵母内核膜蛋白Bqt4具有双链DNA的结合能力,诱导了端粒被招募到细胞核内膜上的起始(Priming)过程(Structure 2019)。在此基础上,核膜蛋白Bqt4通过和端粒蛋白Rap1的相互作用将端粒定位到核膜上。复合物结构分析和生化分析显示端粒蛋白Rap1的N端和C端的丝氨酸可以被不同的激酶磷酸化,分别增强和减弱Bqt4-Rap1之间的相互作用,从而使得端粒和核膜的连接呈现出细胞周期性调控模式(Nucleic Acids Res 2019)。
https://academic.oup.com/nar/article/47/3/1573/5193554
https://www.cell.com/structure/fulltext/S0969-2126(18)30371-X
揭示了端粒蛋白复合物的结构和功能
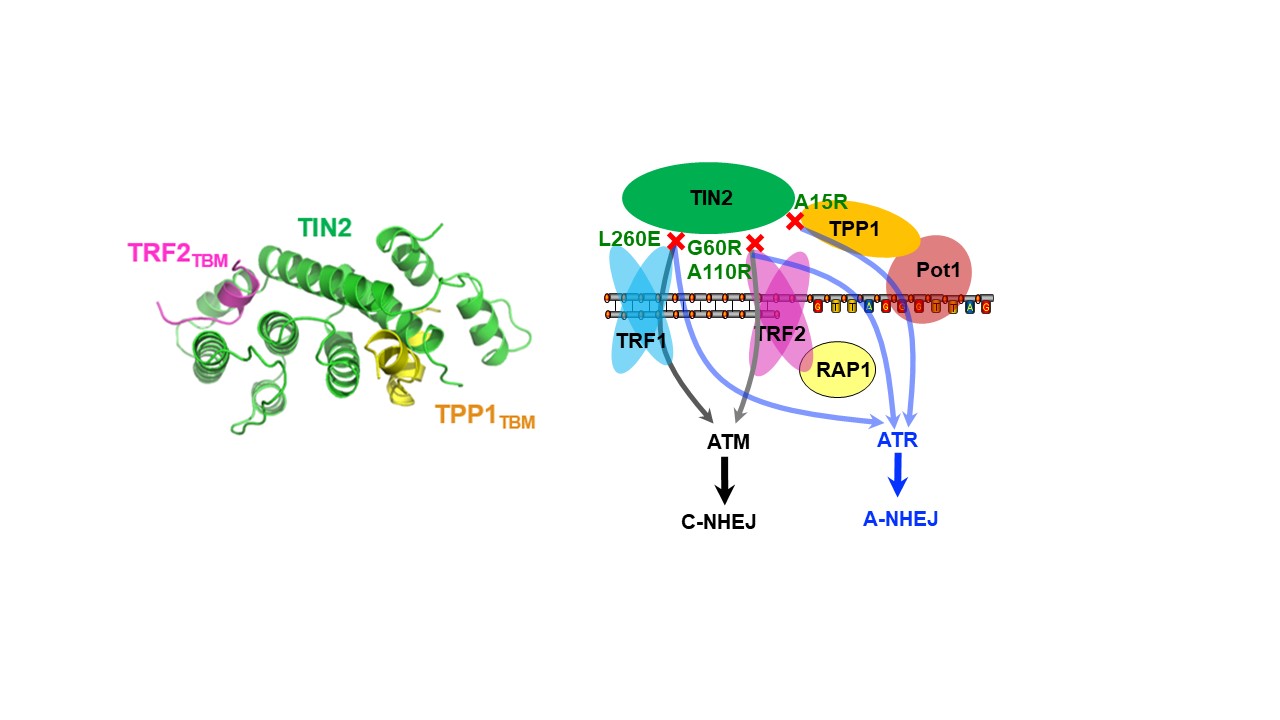
端粒结合蛋白在哺乳动物细胞中包括TRF1、TRF2、POT1、TPP1、TIN2及RAP1这六个核心组分,它们共同组成一个保护性复合物(Shelterin),来调节端粒长度稳态和避免错误的DNA损伤修复。实验室过去解析了多个Shelterin亚复合物的结构,包括TRF1-TIN2,TRF2-TIN2, RAP1-TRF2等(Science 2008; NSMB 2012)。近期我们通过解析端粒Shelterin复合物中一个重要的“桥梁”复合物TIN2-TPP1-TRF2的晶体结构,揭示了TIN2同时结合TRF2和TPP1介导端粒复合物协同组装的作用机制,确定了TIN2,TRF1,TRF2,TPP1等蛋白在抑制ATM和ATR损伤信号、抑制NHEJ(非同源末端连接)和HR(同源重组)途径中的精确功能(Cell Research 2017)。
https://www.nature.com/articles/cr2017144